The Retrosplenial Cortex: Where Landmarks Find Their Place on the Map
The retrosplenial cortex (RSC) produces a distinctive navigation deficit: patients recognize every landmark but cannot use any of them to find their way. This article covers RSC anatomy (Brodmann areas 29/30 behind the corpus callosum), the landmark paradox from Maguire's 2001 lesion review, Epstein's 2008 PPA/RSC dissociation showing RSC tracks spatial knowledge of familiar places, the reference-frame translation hypothesis, Park & Chun's 2009 panoramic fMRI result, and why RSC appears in the default mode network alongside spatial memory and future planning.

The retrosplenial cortex (RSC) produces one of the strangest navigation failures ever recorded in clinical neurology. Patients with damage to this small region of medial parietal cortex can walk into a room, correctly name every object in it, recognize a familiar face across the street, and know that they live in Boston. Then they step outside, look at a street they have seen a thousand times, and have no idea which way to go.
They see perfectly. Their memory for events is largely intact. They know the landmarks. But the landmarks have lost their navigational meaning — they cannot be used to orient, to situate, or to route. Eleanor Maguire's 2001 review of RSC lesion cases found the same description across every patient: capable of landmark recognition; incapable of landmark-guided navigation. 1 That dissociation has made the RSC one of the most actively studied regions in human spatial cognitive neuroscience.
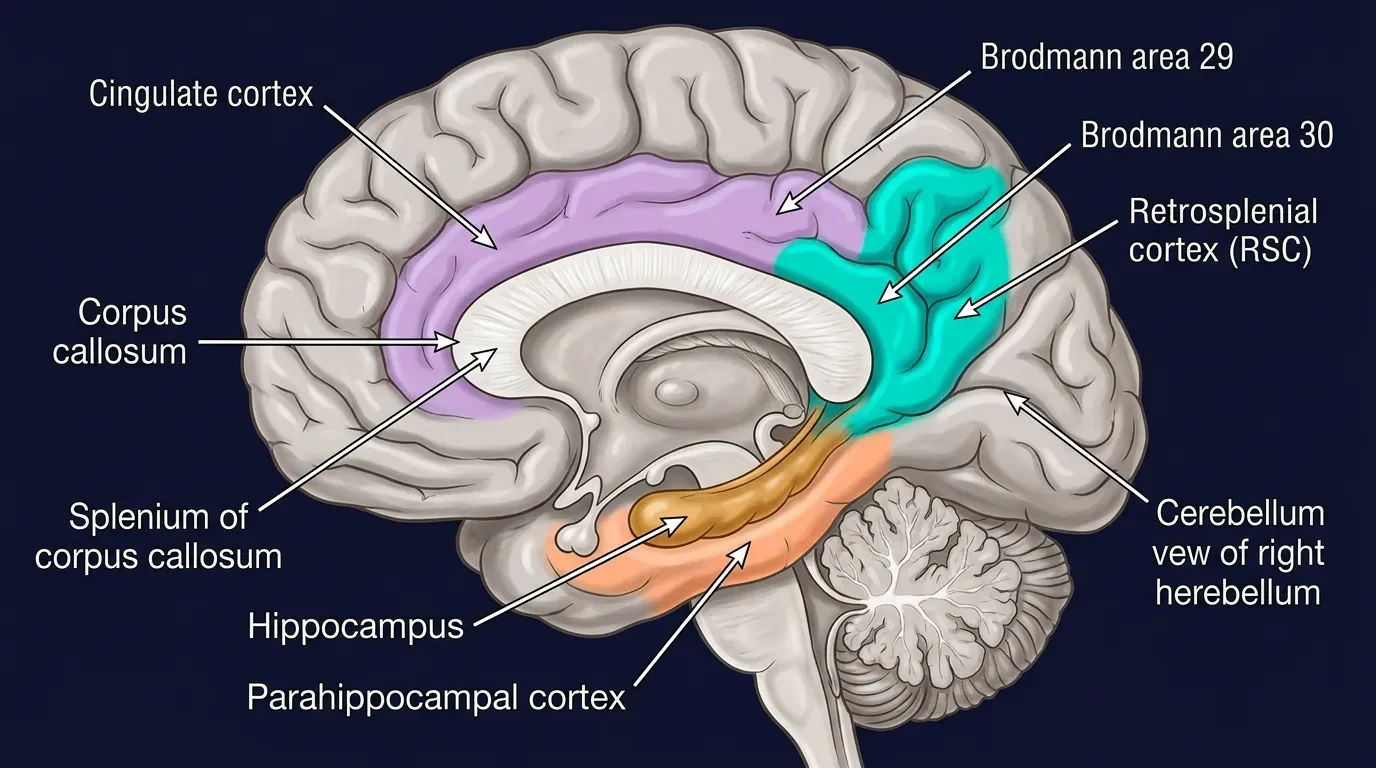
Anatomy: tucked behind the corpus callosum
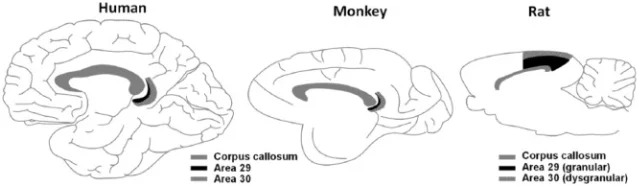
The RSC sits in the posteromedial cortex, immediately behind the splenium of the corpus callosum. Cytoarchitectonically it comprises Brodmann areas 29 (granular) and 30 (dysgranular), which together form a narrow crescent on the medial surface of each hemisphere. In humans the region lies partly within the callosal sulcus and partly on the isthmus of the cingulate gyrus; in rodents it reaches the brain surface more dorsally, making the rat an especially convenient model for recording studies. 2
What makes this address significant is its connectivity. The RSC is wired as an intersection between systems that rarely talk directly: it receives inputs from hippocampal CA1 and the subiculum (the output gate of the hippocampal memory system), connects reciprocally with the anterior thalamic nuclei and the laterodorsal thalamus (major relays for head-direction signals), and projects to and from the parahippocampal region, posterior parietal cortex, and prefrontal cortex. 3 No other region in the posterior cortex sits at the junction of the hippocampal memory circuit, the head-direction signal pathway, and visuospatial cortex all at once.
Scene-selective — but not like the PPA
In the late 1990s, neuroimaging studies of navigation consistently activated two regions in the posterior cortex: the parahippocampal place area (PPA, covered in a previous issue) and a more posteromedial region often called the retrosplenial complex or RSC. Both respond strongly to photographs of scenes and buildings; neither fires much to faces or objects. But they are not doing the same job. 4
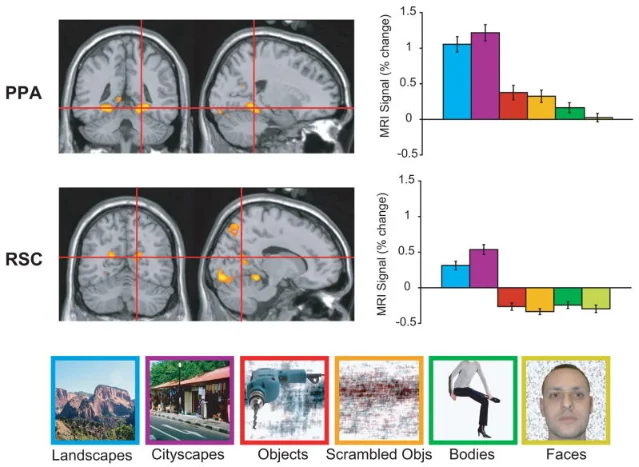
The cleanest dissociation came from Russell Epstein's laboratory at Penn. When participants viewed photographs of their own home campus, RSC responded about 50% more strongly than when they viewed an equally interesting unfamiliar campus — even when task demands were equated. The PPA showed essentially no familiarity effect: it responded the same to Penn and to a random foreign university. 4 The PPA, it seems, is extracting the visual geometry of whichever scene is in front of it; the RSC is also reaching into long-term memory to retrieve what it knows about where this place sits in the broader world.
The campus experiment: what RSC actually retrieves
Epstein's clearest test used Penn students as participants. While they lay in the scanner, photographs of their home campus were shown alongside photographs of an unfamiliar one. For each image, the task alternated: sometimes the students had to retrieve the location or their orientation within the scene (spatial retrieval); sometimes they just had to judge familiarity. 4
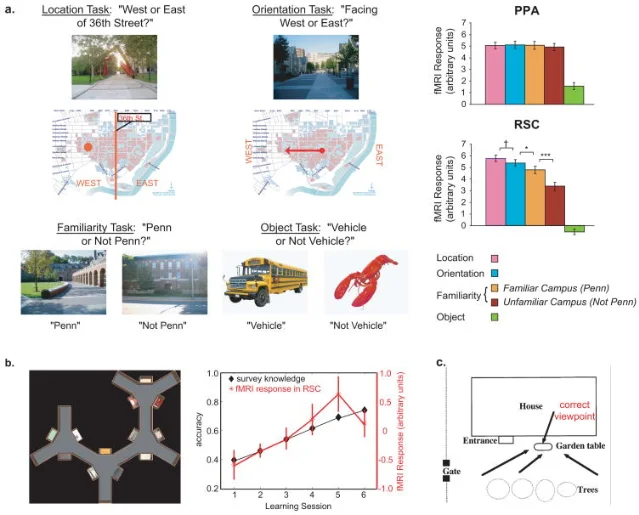
RSC response was highest during the spatial tasks on the familiar campus — highest precisely when the participant was mentally situating themselves in a known map. A companion finding reported that RSC activity during learning of a virtual-reality town tracked the participants' accumulating survey knowledge: as their cognitive map became richer, RSC engagement grew in lockstep. 4 The region is not only activated when navigating; it is building the navigational knowledge base while navigation is learned.
Two reference frames, one translator
A persistent theoretical puzzle in spatial cognition is how the brain gets from the egocentric viewpoint (I'm looking left, the café is behind me) to the allocentric map (the café is on the northeast corner of the block). Every act of navigation requires crossing this translation. The hippocampus maintains an allocentric, world-centered representation — it doesn't care which direction you're facing. Parietal cortex, by contrast, carries egocentric, body-centered spatial codes. Where do these two representations get converted? 2
The RSC is positioned, anatomically and functionally, to be exactly this converter. Its connections reach both parietal cortex and the hippocampal formation; it receives head-direction signals from the anterior thalamic nuclei (which track body orientation continuously). The proposal, supported by several lines of evidence, is that the RSC translates between the two frames of reference: taking the current egocentric view and grounding it in the allocentric map, and vice versa. 4
This hypothesis explains the lesion paradox neatly. After RSC damage, scene-recognition ability is preserved because the PPA's viewpoint-specific visual processing is intact. Landmark knowledge is preserved because object recognition and semantic memory are untouched. What fails is the step that requires translating a recognized landmark into a position and heading within the allocentric map. The patients cannot answer the question their navigation system needs answered: I see the café — now which way does that mean I should be facing?
The 2009 panoramic scene study by Park & Chun (MIT/Yale) provided a direct fMRI test of this idea. 5 When participants watched three sequentially overlapping views of the same panoramic scene — mimicking the natural sweep of the eyes around a room — the PPA treated each view as distinct (no repetition suppression). The RSC, by contrast, recognized the three views as aspects of a single place: it showed clear repetition suppression, responding as if the whole panorama had been seen once, not three times. Critically, this integration collapsed when the views were presented out of context (with unrelated images between them): the RSC needs temporal and spatial continuity to stitch views together. This is exactly what a reference-frame translator would do — maintaining a running map of the surrounding scene even as the eyes move.
The RSC and the default mode network
One reason the RSC was initially underestimated as a navigation region is that it appears in the default mode network (DMN) — the set of regions that activate during rest and deactivate during many externally-directed tasks. This made early researchers assume it was doing something "passive" or self-referential. But closer inspection reveals that DMN activation in the RSC represents internally-directed spatial and episodic processing: mentally constructing scenes, simulating future paths, recalling autobiographical events set in places. 2
The RSC is not activated only by navigating through a real environment. It activates equally when imagining a future journey, when recalling a place from childhood, and when planning a route to a meeting. Vann, Aggleton & Maguire's 2009 review in Nature Reviews Neuroscience proposed that this points to a single deeper function: scene construction — the mental process of building a coherent spatial scene in memory. 2 Navigation, episodic memory, and future planning all require it.
This also explains why the RSC is one of the first regions to show metabolic changes in the progression of Alzheimer's disease — often years before clinical symptoms appear. 3 A patient losing RSC function loses the ability to situate self in place, which manifests early as topographic disorientation and later contributes to the profound spatial memory loss that characterizes the disease.
What remains debated
Three questions sit at the center of current RSC research, as laid out in Mitchell et al.'s 2018 review: 3
Landmark stability vs. reference-frame translation vs. memory consolidation. The RSC responds more to stable landmarks (a mountain, a permanent building) than to transient ones (a parked car). One interpretation is that it tracks which features of the environment are reliable anchors for a cognitive map. Another interpretation focuses on the reference-frame translation role described above. A third assigns the RSC a time-limited role in consolidating hippocampal spatial memories into neocortex for long-term storage. All three may be partly correct — the RSC may serve all these roles, with different subregions or circuits doing each.
Where exactly in the human cortex is the RSC? There is genuine anatomical ambiguity. The fMRI literature has applied "RSC" and "retrosplenial complex" to slightly different patches of cortex across studies; the anatomically precise RSC (areas 29/30 in the callosal sulcus) may be smaller than the "RSC" typically identified with scene-contrast localizers, which often includes the adjacent medial parietal area sometimes called the medial place area. 6
How is the RSC related to the broader scene-processing triad? The current consensus — three scene-selective regions (OPA, PPA, RSC) — assigns each a distinct computation: OPA for local boundary detection and locomotion guidance (covered in the last issue), PPA for viewpoint-specific scene geometry encoding, and RSC for memory-based situating of scenes within a broader spatial framework. How these three regions collaborate in real-time navigation, and whether other regions (the parahippocampal cortex proper, the angular gyrus) need to be added to the picture, remains active territory.
Why the RSC matters
The three scene areas studied in the previous issues of this channel — the OFA, then the body areas, then the OPA — each illustrated what functional specialization looks like at the level of a single cortical patch. The RSC extends this picture in a different direction: it is where perception meets memory, where the currently visible scene docks into the remembered world.
Strip out the RSC and a person navigates like someone looking at a map with no GPS signal and no memory of how they arrived at their current location. They can read the map, recognize the features, recall that London exists — but cannot answer: where am I on this map, right now, given what I see? That is what the retrosplenial cortex computes, drawing on head-direction signals, hippocampal memory, and the visual scene simultaneously, to produce the continuous, grounded spatial self-knowledge that makes purposeful movement through the world possible.
Landmark paper: Vann SD, Aggleton JP, Maguire EA (2009). What does the retrosplenial cortex do? Nature Reviews Neuroscience, 10(11), 792–802. doi:10.1038/nrn2733 2
Course connection: This concept maps to MIT 9.13 (The Human Brain) coverage of scene-selective regions and spatial navigation circuits — the RSC completes the triad of scene-selective areas (OPA, PPA, RSC) and connects scene perception to the hippocampal navigation system discussed in the navigation and memory modules.
참고 출처
- 1Maguire 2001 Neuropsychologia — RSC lesion review
- 2Vann, Aggleton & Maguire 2009 Nat Rev Neurosci — What does the RSC do?
- 3Mitchell et al. 2018 Brain & Neuroscience Advances — RSC and spatial cognition
- 4Epstein 2008 Trends Cogn Sci — PPA and RSC contributions to navigation
- 5Park & Chun 2009 NeuroImage — PPA and RSC in panoramic scene perception
- 6Dilks et al. 2022 Trends in Cognitive Sciences — three cortical scene systems
이 콘텐츠를 둘러싼 관점이나 맥락을 계속 보강해 보세요.